大規(guī)模金屬礦藏的開(kāi)采,產(chǎn)生了大量硫化金屬尾礦如黃鐵礦、砷黃鐵礦等. 這些尾礦給環(huán)境帶來(lái)了很大威脅,如長(zhǎng)期暴露于空氣中,則會(huì)與微生物和水發(fā)生風(fēng)化、 溶浸、 氧化和水解一系列反應(yīng)形成酸性礦山廢水(acid mine drainage,AMD)[1]. 酸性礦山廢水pH非常低且含有大量金屬離子,如鐵、 銅、 鋅、 錳等,對(duì)周?chē)w、 土壤環(huán)境帶來(lái)了十分嚴(yán)重的危害[2,3]. 在黃鐵礦等硫化礦物的氧化過(guò)程中,Leptospirillum ferrooxidans、 Acidithiobacillus ferrooxidans和Ferroplasma spp.等嗜酸微生物發(fā)揮著重要的催化作用[4]. 有嗜酸微生物參與時(shí),黃鐵礦的氧化速率明顯高于化學(xué)氧化對(duì)黃鐵礦的氧化作用,在適宜條件下它們可使整個(gè)反應(yīng)的速度提高106倍[5].
低pH和高金屬含量不僅影響周?chē)貐^(qū)的生態(tài)系統(tǒng),而且影響元素的生物地球化學(xué)循環(huán). 氮循環(huán)是元素地球化學(xué)循環(huán)中非常重要的一部分. 氮循環(huán)主要由固氮、 氨化、 硝化、 反硝化等過(guò)程組成,均由微生物驅(qū)動(dòng). 氨氧化過(guò)程不僅是硝化作用的限速步驟,更是整個(gè)氮元素循環(huán)的中心環(huán)節(jié)[6,7]. 在很長(zhǎng)一段時(shí)間里,氨氧化細(xì)菌(ammonia-oxidizing bacteria,AOB)被認(rèn)為是唯一可以催化氨氧化的微生物[8],其存在直接影響硝化作用強(qiáng)度以及硝化速率[9]. 但近幾年的研究卻發(fā)現(xiàn),氨氧化古菌(ammonia-oxidizing archaea,AOA)廣泛地分布在海洋、 土壤、 沉積物、 淡水以及污水處理廠(chǎng)等諸多環(huán)境中[10],而且在大多數(shù)土壤中AOA是主要的氨氧化承擔(dān)者[11]. 已有研究表明,在pH 3.7~6.0的酸性紅壤中,氨氧化古菌(AOA)的氨單加氧酶α亞基基因(amoA)拷貝數(shù)多于細(xì)菌amoA[11],表明氨氧化古菌可能發(fā)揮著更重要的作用. 但關(guān)于pH小于3的極端酸性環(huán)境中氨氧化類(lèi)群的研究,目前還鮮見(jiàn)報(bào)道.
本研究取樣點(diǎn)位于安徽省某酸性礦山廢水區(qū)域,采集有短小植被覆蓋區(qū)域的根際土壤,采用分子生物學(xué)技術(shù)分析其細(xì)菌和古菌的組成結(jié)構(gòu)并探討承擔(dān)氨氧化功能的微生物類(lèi)群. 研究結(jié)果不但有助于人們?cè)黾訉?duì)礦區(qū)土壤微生物組成的了解,而且擴(kuò)展了人們對(duì)極端酸性土壤中氮循環(huán)機(jī)制的認(rèn)識(shí),有利于對(duì)污染土壤的生物治理和生態(tài)恢復(fù).
1 材料與方法
1.1 樣品采集及分析
樣品于2012年8月采集于安徽某鐵礦酸性礦山廢水庫(kù)邊有短小植被覆蓋區(qū)域. 采集根際土壤樣品后,裝于無(wú)菌封口袋中,4℃保存運(yùn)輸至實(shí)驗(yàn)室并盡快進(jìn)行實(shí)驗(yàn)操作. 取100 g土壤研磨過(guò)1 mm篩,以去除植物根系; 將50 g過(guò)篩的土壤50℃烘干24 h,計(jì)算含水率. 用KCl溶液作為介質(zhì)測(cè)定樣品pH. 總有機(jī)碳(total organic carbon,TOC)含量由總有機(jī)碳測(cè)定分析儀Multi N/C 2100測(cè)定. 氨氮和硝氮含量采用氯化鉀溶液提取-分光光度法測(cè)定[12]. 使用M3浸提劑[13]制備浸提液,用于有效磷和有效金屬的測(cè)定. 使用經(jīng)離心后的浸提液,采用鉬銻抗分光光度法測(cè)定有效磷含量[14]. 有效金屬含量采用ICP(iCAP 6000,Thermo)測(cè)定,具體測(cè)定方法見(jiàn)文獻(xiàn)[15]. 土壤中微生物豐度(Microbial abundance)采用吖啶橙染色直接計(jì)數(shù)法(acridine orange direct counting,AODC)測(cè)定,具體方法見(jiàn)文獻(xiàn)[16].
1.2 土壤基因組DNA的提取
取0.5 g土壤樣品,用基因組DNA提取試劑盒直接提取樣品中的總DNA(參照MoBio試劑盒生產(chǎn)商建議步驟). 提取出來(lái)的基因組總DNA經(jīng)Nanodrop測(cè)定DNA濃度后置于-20℃保存.
1.3 細(xì)菌16S rRNA基因的擴(kuò)增
利用細(xì)菌通用引物27F(5′-AGA GTT TGA TCM TGG CTC AG-3′)和1492R(5′-TAC GGY TAC CTT GTT ACG ACT T-3′)[17],以土壤基因組DNA為模板,擴(kuò)增樣品中細(xì)菌的16S rDNA基因片段. PCR反應(yīng)條件設(shè)置為預(yù)變性95℃ 5 min,變性94℃ 30 s,退火53℃ 30 s,延伸72℃ 2 min,35個(gè)循環(huán)后,72℃延伸7 min. 所有PCR產(chǎn)物都在-20℃保存.
1.4 古菌16S rRNA基因的擴(kuò)增
以土壤基因組DNA為模板,利用古菌通用引物109F(ACK GCT CAG TAA CAC GT)和934R(GTG CTC CCC CGC CAA TTC CT)[18]擴(kuò)增樣品中古細(xì)菌的16S rRNA基因片段. PCR反應(yīng)條件設(shè)置為預(yù)變性95℃ 5 min,變性94℃ 30 s,退火48℃ 30 s,延伸72℃ 2 min,35個(gè)循環(huán)后,72℃延伸5 min.
1.5 氨單加氧酶基因(amoA)的擴(kuò)增
amoA是氨氧化功能酶——氨單加氧酶(ammonia monooxygenase,AMO)α亞基的編碼基因. 以土壤基因組DNA模板,分別利用氨氧化細(xì)菌amoA引物amoA-1F(GGG GHT TYT ACT GGT GGT)和amoA-2R(CCC CTC KGS AAA GCC TTC TTC)[19]、 amoA-3F(GGT GAG TGG GYT AAC MG)和amoA-4R(GCT AGC CAC TTT CTG G)[20],氨氧化古菌amoA引物AF(STA ATG GTC TGG CTT AGA)和AR(GCG GCC ATC CAT CTG TAT GT)[21]擴(kuò)增樣品中氨氧化細(xì)菌和氨氧化古菌的氨單加氧酶基因片段. 氨氧化細(xì)菌PCR反應(yīng)條件設(shè)置為95℃ 5 min,變性94℃ 30 s,退火48℃ 45 s,延伸72℃ 2 min,35個(gè)循環(huán)后,72℃延伸10 min. 氨氧化古菌PCR反應(yīng)條件設(shè)置為95℃ 5 min,變性94℃ 30 s,退火48℃ 60 s,延伸72℃ 2 min,35個(gè)循環(huán)后,72℃延伸10 min.
1.6 克隆文庫(kù)的構(gòu)建
分別構(gòu)建土壤樣品中細(xì)菌和古細(xì)菌的16S rDNA克隆文庫(kù)以及氨氧化古菌的amoA克隆文庫(kù). 氨氧化細(xì)菌amoA基因經(jīng)反復(fù)擴(kuò)增,均未得到PCR產(chǎn)物,故未構(gòu)建該文庫(kù). 克隆文庫(kù)構(gòu)建方法為:將PCR產(chǎn)物連接到Promega公司的pGEM-T克隆載體上,按照生產(chǎn)商的說(shuō)明將質(zhì)粒轉(zhuǎn)化于E. coli DH5α感受態(tài)細(xì)胞中,涂布于含有Amp/X-gal/IPTG的LB平板,于37℃靜置培養(yǎng)17 h,之后在4℃靜置2 h顯色,隨機(jī)挑取白色菌落,重新培養(yǎng). 利用載體特異性引物對(duì)M13-M4(GTT TTC CCA GTC ACG AC)和M13-RV(CAG GAA ACA GCT ATG AC)進(jìn)行PCR擴(kuò)增驗(yàn)證插入片段大小以篩選陽(yáng)性克隆. 細(xì)菌、 古菌和氨氧化古菌克隆文庫(kù)分別挑取105、 38和43個(gè)陽(yáng)性克隆送交上海生工進(jìn)行測(cè)序.
1.7 序列測(cè)定以及系統(tǒng)發(fā)育分析
利用Bellerophon程序檢測(cè)并去除嵌合體[22]. 然后用Dotur軟件對(duì)所得序列進(jìn)行分類(lèi),計(jì)算香農(nóng)指數(shù)(Shannon)和辛普森指數(shù)(Simpson)并制作飽和曲線(xiàn). 最后選取代表克隆運(yùn)用Blast程序與GenBank+EMBL+DDBJ+PDB中進(jìn)行相似性搜索并下載相似性最高的序列和相似性較高的已知種屬的序列. 將所有序列用BioEdit中的ClustalW程序進(jìn)行比對(duì),并用MEGA 4.0軟件構(gòu)建系統(tǒng)發(fā)育樹(shù).
1.8 序列登錄號(hào)
將本研究中所得序列提交至GenBank中,細(xì)菌16S rDNA的序列登錄號(hào)為KF863913-KF863952,KF924200-KF924205; 古菌16S rDNA的序列登錄號(hào)為KF918278-KF918306; 氨氧化古菌amoA的序列登錄號(hào)為KF916687-KF916696.
2 結(jié)果與分析
2.1 AODC結(jié)果與理化分析
所采集土壤樣品的部分物理化學(xué)參數(shù)見(jiàn)表 1. 樣品酸性很強(qiáng),pHKCl僅為2.83. 總有機(jī)碳含量為0.675%,低于農(nóng)田和森林土壤TOC平均含量[23,24]; 含水率為9.5%; 土壤微生物豐度為3.1×108 個(gè) ·g-1; NH+4-N和NO-3-N含量分別為7.6 mg ·kg-1、 3.1 mg ·kg-1. 經(jīng)測(cè)得樣品中有效金屬含量很高,其中Al含量最高達(dá)1 380.3 mg ·kg-1,F(xiàn)e含量高達(dá)57.0 mg ·kg-1. SO2-4含量高達(dá)2 168.0 mg ·kg-1.
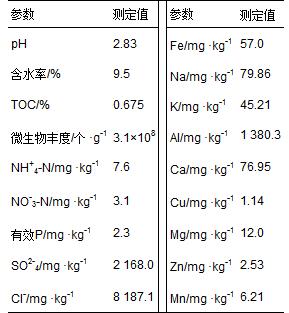
表 1 土壤樣品的部分化學(xué)參數(shù)
2.2 飽和度及多樣性指數(shù)分析
經(jīng)測(cè)序細(xì)菌文庫(kù)共獲得103條序列,去除載體和嵌合體后有效序列為95條; 古細(xì)菌與氨氧化古菌文庫(kù)分別得到28條和39條有效序列. 3個(gè)文庫(kù)的飽和曲線(xiàn)如圖 1所示,3個(gè)文庫(kù)飽和曲線(xiàn)均趨于平緩,可以比較完整地反映樣品中細(xì)菌、 古菌和氨氧化古菌群落結(jié)構(gòu). 從香農(nóng)指數(shù)(Shannon)和辛普森指數(shù)(Simpson)可以看出該樣品中細(xì)菌的多樣性高于古菌(表 2).

圖1 細(xì)菌、 古菌和氨氧化古菌飽和度曲線(xiàn)
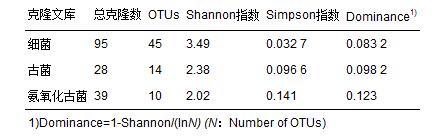
表 2 細(xì)菌、 古菌以及氨氧化古菌克隆文庫(kù)多樣性指數(shù)
2.3 土壤中細(xì)菌、 古菌以及氨氧化古菌多樣性以及系統(tǒng)發(fā)育學(xué)分析 2.3.1 細(xì)菌多樣性以及系統(tǒng)發(fā)育分析
將每種基因型的序列輸入RDP網(wǎng)站,得到該基因型所屬的系統(tǒng)發(fā)育類(lèi)群. 結(jié)果顯示,細(xì)菌文庫(kù)中包含了11個(gè)類(lèi)群(如圖 2),其中所占比例最大的3個(gè)類(lèi)群分別為Acidobacteria(47.4%)、 Verrucomicrobia(18.9%)、 Chloroflexi(10.5%).

圖 2 細(xì)菌文庫(kù)中各類(lèi)群所占比例
經(jīng)NCBI數(shù)據(jù)庫(kù)中Blast程序比對(duì)分析后,選取主要克隆進(jìn)行比對(duì),發(fā)現(xiàn)與該礦區(qū)土壤樣品中細(xì)菌親緣關(guān)系較近的克隆均來(lái)自于一些酸性環(huán)境中,如某些礦區(qū)酸性土壤、 酸性濕地底泥、 酸性紅壤和酸性礦山廢水等,或者植被根際等.
本研究中酸桿菌門(mén)是所占比例最高的門(mén)類(lèi),在文庫(kù)中豐度達(dá)47.4%. 酸桿菌門(mén)細(xì)菌在環(huán)境中分布廣泛,尤其在土壤和沉積物中所占比例較高——在細(xì)菌16S rRNA基因文庫(kù)中的豐度可達(dá)10%~50%[25, 26, 27, 28, 29, 30]. 該樣品中酸桿菌門(mén)細(xì)菌多屬于Gp1類(lèi)群(如圖 3). 酸桿菌門(mén)的一些成員對(duì)于極端環(huán)境如金屬污染和酸性環(huán)境具有耐受性[26].Lee等[29]在研究韓國(guó)某地栗子樹(shù)根際土壤時(shí),分別應(yīng)用PCR以及反轉(zhuǎn)錄PCR(reverse transcription PCR)研究土壤樣品中微生物多樣性,在rRNA基因文庫(kù)以及由rRNA反轉(zhuǎn)錄得到的cDNA文庫(kù)中,酸桿菌門(mén)所占比例都大于50%,表明酸桿菌門(mén)細(xì)菌在該土壤樣品中數(shù)量最多而且新陳代謝活性最高.

圖 3 基于16S rDNA序列的Acidobacteria細(xì)菌系統(tǒng)發(fā)育樹(shù)
克隆Bac-XC-105(2.0%)、 Bac-XC-109(5.1%)、 Bac-XC-19(1.0%)和Bac-XC-25(1.0%)均與Candidatus Koribacter versatilis 的親緣關(guān)系較近. Ward等[31]比較了包括Candidatus Koribacter versatilis在內(nèi)的酸桿菌門(mén)中3個(gè)種的全基因組,發(fā)現(xiàn)Candidatus Koribacter versatilis具備硝酸鹽和亞硝酸鹽還原能力,且適宜在pH 4.0~6.5的環(huán)境中生存.
克隆Bac-XC-27(4.0%)、 Bac-XC-86(1.0%)和Bac-XC-99(1.0%)與菌株Acidobacteria sp. isolate N3B的16S rDNA序列相似性均為97%,該菌株分離自法國(guó)加爾省一流經(jīng)古代礦區(qū)的酸性溪流中[32]. Acidobacteria屬菌株多為分離自酸性環(huán)境的化能異養(yǎng)菌,革蘭氏染色呈陰性,好氧; 可在pH 3.0~6.0、 溫度為20~37℃的環(huán)境中生存[33].
克隆Bac-XC-14和Bac-XC-4(豐度分別為4.0%和3.0%)與Holophaga sp.的16S rDNA序列同源性較高. Holophaga屬細(xì)菌在一些酸性礦坑水中曾檢測(cè)到,如江西德興市銀山鉛鋅礦的露天礦坑儲(chǔ)水池和地下300多m隧道中自然氧化形成的酸性礦坑水[34]. Holophaga屬細(xì)菌多為革蘭氏陰性嚴(yán)格厭氧菌,可在溫度為10~35℃環(huán)境中生存,最適溫度為28~32℃,最適pH為6.8~7.5[35].
克隆Bac-XC-41(5.1%)和Bac-XC-80(2.0%)在系統(tǒng)發(fā)育樹(shù)上與Acidopila rosea聚為一類(lèi)(圖 3). Okamura等[36]分別在日本的一個(gè)AMD處理廠(chǎng)(水樣pH 2.7)和一個(gè)茶葉種植園土壤(pH 4.5)中分離到兩株Acidipila rosea. Acidipila rosea為革蘭氏陰性好氧菌,細(xì)胞呈球形或球桿形,營(yíng)養(yǎng)類(lèi)型為化能有機(jī)營(yíng)養(yǎng)型; 可以在pH 3.0~6.0范圍內(nèi)生存,最適pH為4.5; pH 7.0時(shí)不能生存.
如圖 4所示,放線(xiàn)菌門(mén)包括3種基因型(占文庫(kù)的4.0%)都與Aciditerrimonas ferrireducens在系統(tǒng)發(fā)育樹(shù)上聚為一簇. Aciditerrimonas ferrireducens是一種中度嗜熱的嗜酸菌,具有還原鐵的能力; 革蘭氏染色呈陽(yáng)性,可以在pH 2.0~4.5,溫度35~58℃的環(huán)境中生存,最適pH為3.0,最適生長(zhǎng)溫度為50℃. 有氧條件下?tīng)I(yíng)養(yǎng)類(lèi)型為化能異養(yǎng),無(wú)氧環(huán)境中為化能自養(yǎng),利用三價(jià)鐵氧化氫獲得能量[37].
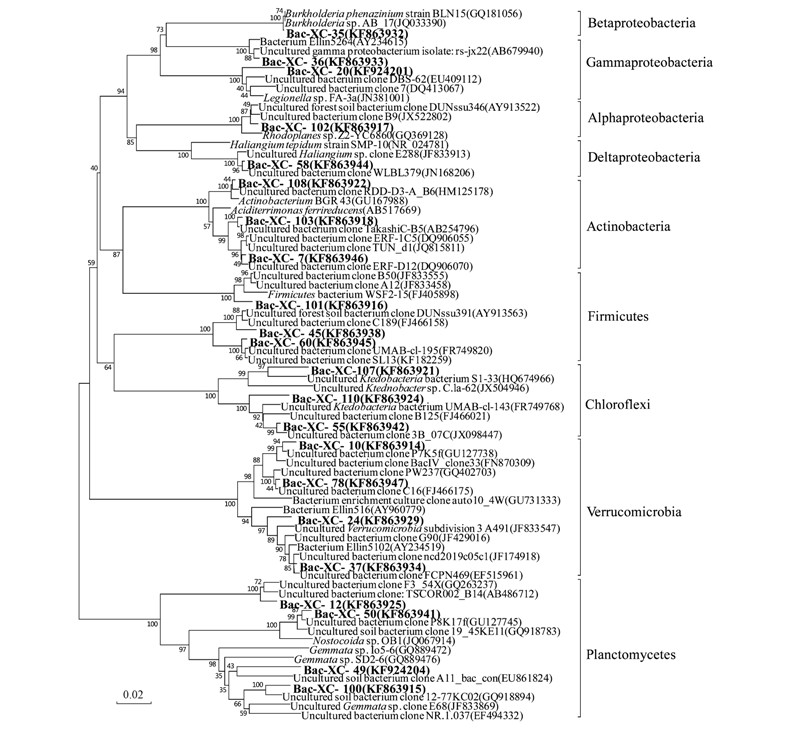
圖 4 基于16S rDNA序列的其它門(mén)細(xì)菌系統(tǒng)發(fā)育樹(shù)
α-變形菌門(mén)在本文庫(kù)中所占比例僅為6.0%. 其中克隆Bac-XC-102(3.0%)與紅游動(dòng)菌屬(Rhodoplanes sp.)Z2-YC6860在系統(tǒng)發(fā)育樹(shù)上聚為一簇. 紅游動(dòng)菌屬細(xì)菌為一類(lèi)紫色非硫細(xì)菌; 細(xì)胞革蘭氏染色呈陰性,兼性厭氧光能營(yíng)養(yǎng)型細(xì)菌,細(xì)胞含有類(lèi)胡蘿卜素和菌綠素; 該類(lèi)細(xì)菌在黑暗厭氧條件下可以完成反硝化過(guò)程進(jìn)行脫氮[38].
2.3.2 古菌多樣性以及系統(tǒng)發(fā)育分析
經(jīng)過(guò)RDP網(wǎng)站的比對(duì)分析,本研究的土壤樣品中古菌屬于兩個(gè)類(lèi)群,即廣古菌門(mén)(Euryarchaeota)和奇古菌門(mén)(Thaumarchaeota). 測(cè)試的29個(gè)克隆分屬于14個(gè)操作分類(lèi)單元(operational taxonomic units,OTUs),其中10個(gè)OTUs(22個(gè)克隆)屬于奇古菌門(mén),4個(gè)OTUs(7個(gè)克隆)屬于廣古菌門(mén). 與細(xì)菌相比,古菌多樣性明顯較低. 通過(guò)系統(tǒng)發(fā)育分析發(fā)現(xiàn)該土壤樣品中古菌與氨氧化作用密切相關(guān).
奇古菌門(mén)是2008年從中溫環(huán)境來(lái)源的古菌(溫泉古菌)劃分出來(lái)的一個(gè)新類(lèi)群[39]. 古菌文庫(kù)中歸類(lèi)于奇古菌門(mén)的古細(xì)菌均與氨氧化作用相關(guān). 奇古菌門(mén)的氨氧化古菌分布廣泛,在沉積物、 土壤、 淡水以及海洋環(huán)境中其amoA含量顯著高于氨氧化細(xì)菌[40]. 一些研究運(yùn)用抑制劑抑制硝化作用結(jié)合使用穩(wěn)定性同位素探測(cè)技術(shù)(stable isotope probing,SIP)等證明了氨氧化古菌在一些自然環(huán)境尤其是寡營(yíng)養(yǎng)環(huán)境的硝化作用中起重要作用[13, 39, 40].
奇古菌門(mén)中比例最高也是文庫(kù)中豐度最大的基因型為Arch-XC-25,所占比例為27.6%. 在GenBank數(shù)據(jù)庫(kù)的比對(duì)結(jié)果中,與該克隆親緣關(guān)系較近的均為未培養(yǎng)菌株(如圖 5),多數(shù)檢測(cè)自土壤和沉積物中; 其中相似度為99%的一個(gè)序列分離自美國(guó)橡樹(shù)嶺Melton河支流流域沿岸pH為4.5的土壤樣品. 克隆Arc-XC-13在文庫(kù)中豐度為13.8%,與該克隆親緣關(guān)系最近的克隆來(lái)自金屬礦區(qū)(如鈾尾礦)和酸性泉水. 基因型Arc-XC-11和Arc-XC-27各包含兩個(gè)克隆(豐度為6.9%),其余基因型均為單克隆. 與這些基因型相似的序列多分離自酸性紅壤、 酸性礦山廢水、 酸性溫泉以及植物根際土壤等. 在GenBank數(shù)據(jù)庫(kù)中與本研究中所有古菌基因型相似度在93%以上的都為未得到純培養(yǎng)的古菌的16S rDNA序列. 而去除未培養(yǎng)古菌的序列后,本研究中所有古菌的序列類(lèi)型都與Nitrososphaera(亞硝化球菌屬)親緣關(guān)系最近,相似度為81%~87%. Nitrososphaera 屬的Nitrososphaera viennensis EN76(維也納亞硝化球菌)是第一株分離自土壤環(huán)境的氨氧化古菌[41]; Nitrososphaera gargensis(加爾加亞硝化球菌)分離自俄羅斯加爾加地區(qū)的一個(gè)溫泉,具有氧化氨的能力,但是氨濃度超過(guò)3 mmol ·L-1會(huì)抑制其生長(zhǎng)[42]. 分離自英格蘭酸性土壤(pH 4.5)的Nitrosotalea devanaterra(阿伯丁土壤亞硝化細(xì)桿菌)是1株嗜酸氨氧化古菌,pH 4~5時(shí)均生長(zhǎng)良好,當(dāng)pH>5.5時(shí)生長(zhǎng)會(huì)受到抑制; 最適生長(zhǎng)溫度為25℃[43]. 已有研究表明,在酸性土壤中,AOA與硝化潛勢(shì)存在明顯正相關(guān)關(guān)系,主導(dǎo)氨氧化過(guò)程的發(fā)生[43]. 本研究土壤也具有類(lèi)似特點(diǎn),氨氧化過(guò)程可能主要由氨氧化古菌驅(qū)動(dòng).
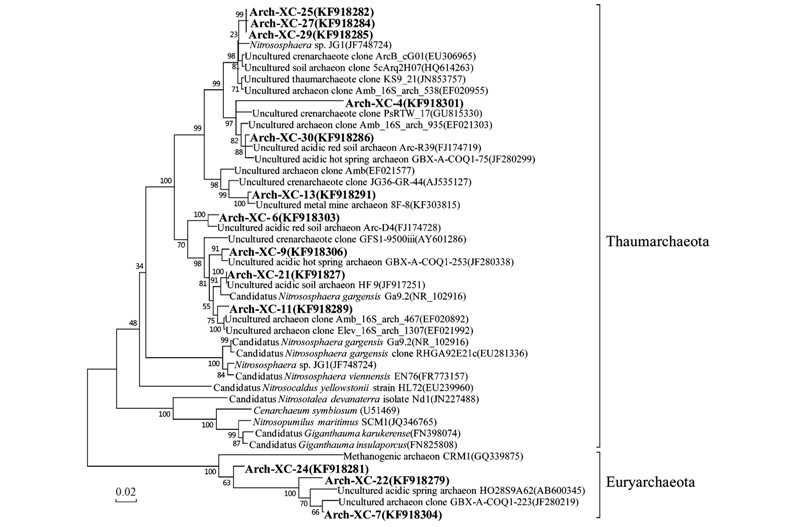
圖 5 基于16S rDNA序列的古菌系統(tǒng)發(fā)育樹(shù)
本研究的文庫(kù)中廣古菌門(mén)包括3種基因型,其中基因型Arc-XC-22豐度最大,為13.8%. 這3種基因型的最相似序列均分離自哥倫比亞安第斯山脈的一個(gè)酸性溫泉[44]和日本箱根大涌谷的一個(gè)酸性溫泉[45]. 克隆Arc-XC-3與數(shù)據(jù)庫(kù)中序列相似度低于80%,可能是廣古菌門(mén)的新種. 廣古菌門(mén)的所有基因型與在GenBank中最相似的菌種均為產(chǎn)甲烷古菌(Methanogenic archaea),相似度均為82%~84%. 產(chǎn)甲烷古菌是一類(lèi)嚴(yán)格厭氧的僅利用二氧化碳和氫氣、 甲酸鹽、 甲醇、 甲胺、 和/或乙酸鹽進(jìn)行能量代謝并產(chǎn)生甲烷的古菌. 這類(lèi)微生物在全球碳循環(huán)中發(fā)揮重要作用[46].
2.3.3 氨氧化古菌多樣性以及系統(tǒng)發(fā)育分析
該土壤樣品未擴(kuò)增到氨氧化細(xì)菌的amoA基因片段,說(shuō)明樣品中不存在氨氧化細(xì)菌或氨氧化細(xì)菌含量很低.
由古菌文庫(kù)組成可知本樣品中大多數(shù)古菌與氨氧化功能相關(guān),故利用古菌特異性氨單加氧酶引物對(duì)AF和AR擴(kuò)增氨單加氧酶基因序列,共得到39條有效基因序列. 經(jīng)與GenBank數(shù)據(jù)庫(kù)中已有序列比對(duì),發(fā)現(xiàn)該土壤樣品中多數(shù)氨氧化古菌的最相似序列來(lái)自酸性紅壤、 酸性土壤和酸性礦山等與本樣品所取條件相似的環(huán)境中,其中多數(shù)與已知菌種的相似性都低于90%,基因型AOA-XC-17和AOA-XC-19與Nitrosotalea devanaterra(阿伯丁土壤亞硝化細(xì)桿菌)的相似性都為94%. 從氨氧化古菌的系統(tǒng)發(fā)育樹(shù)(圖 6)可看出,Cluster 3中的基因型AOA-XC-1(文庫(kù)中豐度為5.1%),AOA-XC-10(12.8%),AOA-XC-13(10.3%),AOA-XC-28(2.6%),AOA-XC-33(2.6%)與數(shù)據(jù)庫(kù)中的氨氧化古菌序列相似性都較低,與已得到純培養(yǎng)的氨氧化古菌(全部位于Cluster 2中)親緣關(guān)系較遠(yuǎn),證明該土壤樣品中存在A(yíng)OA新類(lèi)群.
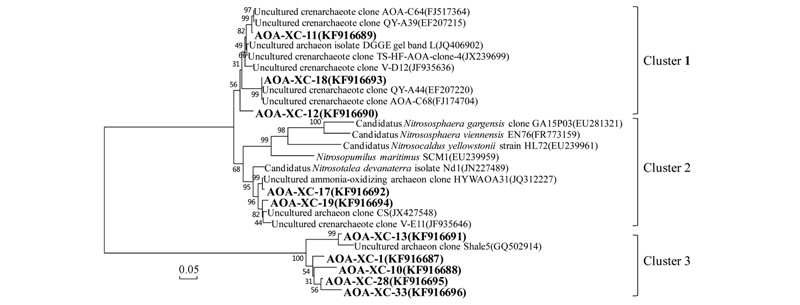
圖 6 基于amoA序列的氨氧化古菌(AOA)系統(tǒng)發(fā)育樹(shù)
3 討論
在前期研究中,筆者利用分子生物學(xué)的方法著重研究了取樣地的酸性礦山廢水庫(kù)的微生物群落結(jié)構(gòu)和該區(qū)域內(nèi)廢礦石中真核生物的多樣性[17,47]. 取樣區(qū)域AMD的pH在3以下[17],周?chē)鷧^(qū)域廢礦石的pH也低于3[47]. 本研究中的土壤樣品受到酸性礦山廢水的嚴(yán)重污染. 該樣品克隆文庫(kù)中細(xì)菌、 古菌序列在數(shù)據(jù)庫(kù)中的最相似序列多分離自酸性土壤、 酸性沼澤、 酸性紅壤以及酸性礦山等理化條件相似的環(huán)境中. 細(xì)菌文庫(kù)中最大的門(mén)類(lèi)為酸桿菌門(mén),在該門(mén)類(lèi)中存在耐酸耐重金屬的類(lèi)群. 此外,文庫(kù)中存在與反硝化作用相關(guān)的菌種,如Candidatus Koribacter versatilis、 紅游動(dòng)菌屬(Rhodoplanes sp.),也存在與鐵還原相關(guān)的菌種,如Aciditerrimonas ferrireducens,但未發(fā)現(xiàn)與氨氧化作用相關(guān)的菌種,說(shuō)明該樣品中不存在或存在較少的氨氧化細(xì)菌. 而在古菌文庫(kù)中卻發(fā)現(xiàn),大部分古菌是與氨氧化作用相關(guān)的奇古菌門(mén)的成員,且與已得到純培養(yǎng)的菌種親緣關(guān)系較遠(yuǎn). Nitrosotalea devanaterra是奇古菌門(mén)中唯一一種嗜酸的氨氧化古菌,但是該細(xì)菌與本研究中所有AOA序列的相似性都極低. 經(jīng)建立古菌amoA克隆文庫(kù),同樣證實(shí)該土壤樣品中存在氨氧化古菌,所以取樣區(qū)域內(nèi)氨氧化過(guò)程主要由氨氧化古菌主導(dǎo). 文庫(kù)中所得到的amoA序列與已得到純培養(yǎng)的氨氧化古菌的amoA序列相似性均低于85%,而且其中5種基因型與數(shù)據(jù)庫(kù)中已有研究的類(lèi)群親緣關(guān)系較遠(yuǎn),在系統(tǒng)發(fā)育樹(shù)上組成一個(gè)獨(dú)立分枝,由此推斷該土壤中可能存在未知的氨氧化古菌類(lèi)群. 礦區(qū)土壤由于其較低的pH和高重金屬含量使其具有特殊性,其中所含微生物對(duì)低pH和重金屬耐受性高,為今后礦區(qū)土壤以及理化性質(zhì)相似的污染土壤的原位生物修復(fù)提供可靠依據(jù).
4 結(jié)論
(1) 取樣區(qū)域土壤受到酸性礦山廢水的嚴(yán)重污染,pH僅為2.83,金屬Fe和Al含量都很高. 文庫(kù)中的序列均與酸性環(huán)境中檢測(cè)到的序列具有較高的同源性,證明該土壤中存在大量耐酸以及嗜酸的微生物.
(2) 細(xì)菌文庫(kù)覆蓋11個(gè)類(lèi)群,酸桿菌門(mén)豐度近50%,第二大門(mén)類(lèi)為疣微菌門(mén),所占比例為18.9%. 古菌文庫(kù)多樣性低,只存在兩大門(mén)類(lèi),分別為奇古菌門(mén)和廣古菌門(mén),前者占優(yōu)勢(shì)地位.
(3) 細(xì)菌文庫(kù)中未發(fā)現(xiàn)與氨氧化作用相關(guān)的類(lèi)群,只存在反硝化細(xì)菌參與N元素的生物地球化學(xué)循環(huán). 該土壤樣品中的氨氧化作用主要由奇古菌門(mén)的氨氧化古菌所驅(qū)動(dòng),且該區(qū)域存在氨氧化古菌的新類(lèi)群.(來(lái)源及其作者:中國(guó)地質(zhì)大學(xué)(北京)水資源與環(huán)境學(xué)院 劉瑩、王麗華、郝春博、李璐、李思遠(yuǎn)、馮傳平)
